
Tamaño texto
Lector contenido

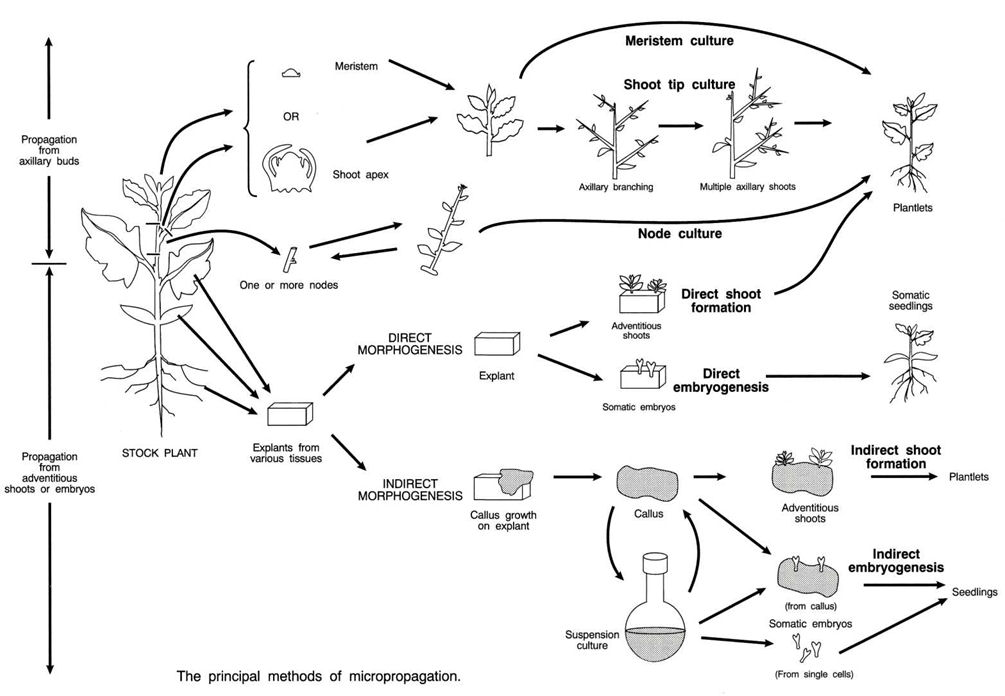
7.2 Fase 1. Iniciación
7.3 Fase 2. Multiplicación
7.3.1 Multiplicación mediante organogénesis adventicia indirecta
7.3.2 Multiplicación mediante organogénesis adventicia directa
7.3.3 Multiplicación mediante yemas preexistentes
7.3.4 Multiplicación mediante embriogénesis
Objetivo: puesta en cultivo de explantos Punto de partida: la toma de un fragmento vegetal hace que éste no reciba sustancias ni estímulos del resto del cuerpo de la planta, por lo que se produce un desequilibrio. Para tener éxito en el cultivo se debe restablecer dicho equilibrio.
Acciones a realizar: adquirir conocimientos sobre la especie y variedad a cultivar (genotipo, etc.), así como de los procedimientos de cultivo in vitro, y específicamente de micropropagación.
La información previa a adquirir es respecto a:
- Selección y localización del individuo concreto a micropropagar
- Estado fisiológico de la planta madre • Conocimiento y optimización del estado sanitario de la planta madre
- Tratamientos específicos previos a realizar, en su caso (ruptura de dormición, termoterapia, prelavados, tratamientos fitosanitarios, aislamiento, etc)
- Composición del medio de cultivo y reguladores
- Selección de las condiciones de cultivo (recipientes, iluminación, temperatura, etc.)
Objetivo: observar la iniciación del desarrollo de los explantos en las condiciones adecuadas, y en el sentido previsto.
Acciones a realizar: siguiendo lo determinado en la fase de preparación, se procederá a la:
- Separación de un fragmento de la planta madre
- Esterilización del material vegetal
- Obtención del explanto a cultivar
- Incubación del explanto en las condiciones preestablecidas
- Espera: tiempo de respuesta
- Observación del éxito de la iniciación: el explanto se desarrolla libre de contaminaciones y en el sentido previamente seleccionado.
Principales problemáticas:
- Contaminaciones
- Falta de desarrollo del explanto
- Desarrollo diferente al esperado o Pardeamiento
Objetivo: Producción de multitud de explantos idénticos genéticamente, con desarrollo homogéneo, capaces de transformarse en plántulas completas.
Acciones a realizar: dejar desarrollarse los explantos durante un tiempo (ej. 3-6 semanas) y subcultivarlos periódicamente para multiplicar su número. Elegir para ello una vía de multiplicación.
Vías de multiplicación: se dispone de varias opciones derivadas de la combinación de tres pares de alternativas, si bien en la realidad no pueden producirse todas las combinaciones. En varios casos es posible multiplicar la planta mediante más de una opción, seleccionándose la que mejores resultados ofrezca. Las vías de multiplicación son:
- Organogénesis vs. Embriogénesis
- Yemas preexistentes vs. Estructuras adventicias
- Directa vs. Indirecta
Gautheret
Es lo que se denomina cultivo de callo. El desarrollo de callo es un crecimiento de tipo tumoral que se produce por un desequilibrio hormonal en las células del explanto. El callo está formado por células vegetales que se dividen rápidamente de forma asíncrona y en el que no existe una programación de la diferenciación, de forma que no pueden producirse órganos ni tejidos con funciones determinadas y no da lugar a plántulas definidas.

Callo de violeta africana
a) Tipos de explantos que originan callo.
Cualquiera, siempre que reciba el estímulo apropiado. Los explantos preferentes son los que cumplen las siguientes características:
- Sin programa de diferenciación predefinido
- Escasa diferenciación
- Alta tasa de división celular
Son explantos habituales para el desarrollo de callo los fragmentos de hojas o de tallo que contengan tejidos vasculares, siempre sin yemas axilares.
b) Tipos de callo: potencial morfogénico y regeneración
Aunque el callo sea un crecimiento desorganizado y de tipo tumoral, no todos los callos son equivalentes. Algunos tienen un alto potencial morfogénico y de regeneración, es decir, recibiendo los estímulos adecuados en un determinado momento pueden iniciar una organización celular y producir estructuras adventicias, yemas vegetativas o embriones somáticos, que se desarrollarán posteriormente como tallos o como embriones enteros. Este tipo de callo morfogénico es el que se busca; una vez se tenga el suficiente número de estos callos morfogénicos se transferirán a otro medio de cultivo que incluya los reguladores de crecimiento adecuados para promover dicha organización celular e iniciar la obtención de las correspondientes plántulas (o en su caso de los embriones).
La morfogénesis de los callos pueden dirigirse en diferentes sentidos; de ellos no es deseable la formación de raíces, y se busca la de tallos adventicios o embriones.

Callo de violeta africana

Callo de zanahoria
c) Eficiencia del cultivo de callo como sistema de multiplicación in vitro.
Depende de:
- Intervalo entre iniciación y regeneración
- Número de subcultivos posibles (hacer aquí un clic al siguiente apartado)
- Formación de estructuras adventicias: tasa y frecuencia
- Capacidad de regeneración de tallos (o embriones)
- Capacidad posterior de desarrollo de raíces para la producción de plántulas completas
Número de subcultivos posibles
Teóricamente un callo se puede subcultivar indefinidamente, sin embargo en la práctica el número recomendable de subcultivos es limitado. El callo se obtiene gracias a que se provoca un desequilibrio celular mediante el aporte de reguladores de crecimiento, este desequilibrio se va incrementando con el tiempo de cultivo en presencia de dichos reguladores, que se acumulan en las células y pueden favorecer la aparición de mutaciones. Si la concentración de reguladores es excesivamente elevada, o si se mantiene durante un tiempo excesivo el porcentaje de explantos mutados será alto; en un determinado número de ellos las mutaciones serán deletéreas y no se podrán producir plántulas completas, reduciendo por tanto la eficiencia del sistema, otro porcentaje producirá plántulas completas pero que lleven mutaciones no deletéreas e indeseables en un sistema de micropropagación.
d) Ventajas y desventajas de la multiplicación vía callo.
Es la más rápida, pero tiene una baja estabilidad genética para la mayoría de las especies, debido a que el desequilibrio para causar el desarrollo del propio callo favorece la aparición de mutaciones; esta inestabilidad genética hace que este procedimiento sólo se utilice en determinadas ocasiones.
e) Metodología
El sistema para inducir la formación de yemas adventicias incluye la adición de reguladores de crecimiento en el medio de cultivo, preponderando las auxinas sobre las citoquininas; en concreto para inducir el desarrollo de callo suele utilizarse el 2,4-D combinado con benciladenina o con quinetina, si bien depende de la especie a cultivar.
La organogénesis adventicia directa consiste en inducir la diferenciación de yemas de tallo directamente sobre un tejido que no las tenga preformadas y sin que éste pase por un estado previo de formación de callo. Como en el cultivo de callo este sistema evita la utilización de yemas preformadas, pero consigue que se formen nuevos primordios foliares donde no los había, induciendo un nuevo patrón de diferenciación.
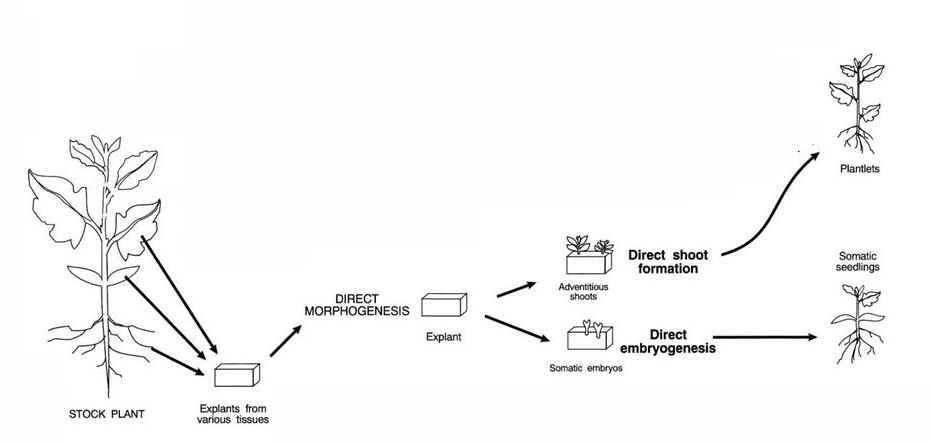
a) Tipos de explatos
Si bien teóricamente es posible la organogénesis adventicia directa a partir de cualquier grupo de células, con algunos exlantos se puede llevar a cabo mas fácilmente, estos son:
- Explantos de hoja. Son los mas frecuentes. Por ejemplo: Begonia, Crasuláceas, Pelargonium, Peperomia, Saintpaulia (un buen número de plantas ornamentales).
- Explantos de zonas internodales del tallo.
- Explantos de bulbo. En las plantas que producen este tipo de estructuras se usan preferentemente fragmentos de hojas del bulbo, por ejemplo: Hyacinthus, Narcissus, Allium, etc.
- Explantos de raíz: mora, frambuesa, algunos manzanos.
b) Eficiencia del cultivo
La tasa de propagación que puede obtenerse subcultivando este tipo de explantos en medios que continuamente induzcan la formación de yemas adventicias, aunque más lenta que en el caso de cultivo de callo, es también muy elevada. Como prueba de ello se ha calculado que la capacidad de multiplicación de Begonia, tomando un fragmento cuadrado de hoja joven de 7cm de lado, es de una producción 1014 plantas al final de un año de subcultivos.
c) Ventajas y desventajas de la multiplicación por organogénesis adventicia directa
La multiplicación mediante formación de yemas adventicias sin formación previa de callo no es tan rápida como la vía de callo, pero sigue siendo muy rápida, por lo que el descenso de velocidad no es una gran desventaja. Su estabilidad genética es sensiblemente mayor que la del callo, si bien existen casos en que se ha observado una proporción excesiva de mutaciones en la descendencia; esto se puede deber a la formación de microcallos en los explantos, los cuales no se podrían observar fácilmente a simple vista debido a la gran profusión del desarrollo del material vegetal. Estos microcallos se forman frecuentemente por un exceso en las dosis de reguladores aplicados. El sistema se puede aplicar en diversos tipos de especies, tal y como se ha indicado en relación al tipo de explantos utilizados, sin embargo no es posible utilizarlo para la propagación de plantas que sean quimeras o que presenten un aspecto variegado debido a la presencia de virus en su interior.
d) Metodología
El sistema para inducir la formación de yemas adventicias incluye la adición de reguladores de crecimiento en el medio de cultivo, preponderando las citoquininas sobre las auxinas. En este caso es recomendable evitar el uso de 2,4-D como auxina, ya que favorece el desarrollo de callo.
Como ejemplo se puede citar la formación de 100 bulbillos al cultivar los fragmentos de una hoja escamosa del bulbo de Lilium durante 4-6 semanas, lo que constituye una tasa de micropropagación rápida. En el extremo contrario un sistema lento es el descrito para Narcissus: una hoja escamosa de su bulbo que se fragmenta en diversos explantos y al cultivarla en un medio con 4-12 mg.l-1 de Benciladenina y 1-2 mg.l-1 de NAA sólo produce unos 20 bulbillos, si bien si se compara con la velocidad de propagación tradicional el avance es sustancial, ya que sólo produciría 2 bulbillos en un tiempo equivalente.
Algunas de las especies a las que se ha aplicado con éxito la multiplicación vía formación de yemas adventicias son: Gesneracias (Achimenes, Sinningia, Saintpaulia, Streptocarpus), Begonias, Gerberas, Cactaceas, y algunas monocotiledóneas que forman bulbos (Hyacinthus, Narcissus, Allium, Lilium).
a) Tipos de explanto
La multiplicación por yemas preexistentes consiste en promover el máximo desarrollo de las yemas que contiene un explanto, por lo tanto debe realizarse mediante una organogénesis directa. Por la misma razón el tipo de explanto a utilizar estará limitado a cualquiera que contenga yemas preexistente: una yema apical de tallo o una sección nodal de tallo con uno o más nudos; también se considera la posibilidad de que el explanto sea un ápice de raíz, aunque este tipo de cultivos es muy infrecuente como sistema de micropropagación.
Este sistema es el más semejante al de la propagación convencional, ya que simplemente utiliza las yemas que ya existen, hace que se desarrollen como tallos y que posteriormente enraícen, dando así lugar a plantas completas. Las diferencias respecto a la propagación in vivo son simplemente el reducido tamaño del explanto, que a partir de un individuo permite obtener un número de explantos mucho más alto que de esquejes, el causar el desarrollo de un gran número de yemas, que en condiciones naturales estarían inactivas, y el facilitar el enraizamiento posterior de todas las plántulas, lo que para muchas especies en condiciones naturales es frecuentemente problemático.
b) Eficiencia del cultivo y metodología
El sistema para inducir la formación de yemas adventicias parte de un explanto con una o varias yemas, que mediante los reguladores adicionados al medio de cultivo (principalmente citoquininas, y una concentración mucho menor de auxinas si fueran requeridas) se desarrolla formando una rama, que a su vez se ramifica; cada una de las ramas nuevas también sufre un máximo de ramificación. En el momento de realizar un subcultivo se fragmenta el explanto en múltiples nuevos explantos, cada uno de los cuales tendrá una o varias yemas que volverán a ramificarse, y así en cada subcultivo. Al finalizar la fase de multiplicación se deberá aislar cada una de las plántulas y cambiarlas a un medio de cultivo en el que ya no se promueva la ramificación sino que se pase a la siguiente fase.
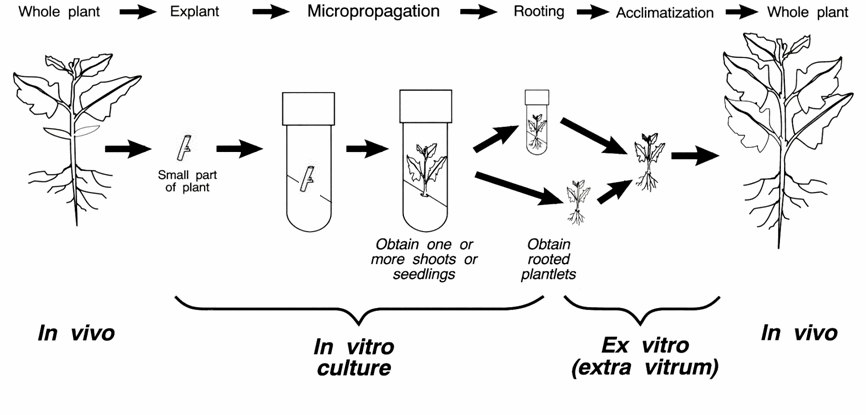
Los explantos suelen subcultivarse como media cada 4-6 semanas, y el número de subcultivos debe limitarse para evitar la aparición de mutaciones por acúmulo de citoquininas o por formación de microcallos que pasen inadvertidos. Una recomendación frecuente es restringir estos subcultivos a 4-5, si bien en función de la especie a micropropagar su número es muy variable.

Cultivos de secciones nodales de pepino (izquierda) y de vid (derecha)
La tasa de multiplicación mediante este procedimiento es logarítmica. El número de plántulas al final de la fase de multiplicación dependerá del número de explantos iniciales (a), del tiempo durante el que se prolongue esta fase de multiplicación y del número de subcultivos que se realicen (n), así como del número de nuevos explantos obtenidos en cada subcultivo (b). Suponiendo que no haya ningún tipo de pérdidas el número de plántulas finales obtenidas será a.bn. Si pusiéramos en cultivo sólo 10 explantos y se subcultivaran mensualmente durante 5 meses, de forma que cada uno de ellos diera lugar a 8 nuevos explantos, al final de esos 5 meses se tendrían más de 32000 plantas, y si se realizara un subcultivo más el número final de plantas superaría las 250000. En la práctica son conocidas algunas especies por su mayor o menor velocidad de multiplicación in vitro mediante este sistema:
- Multiplicación rápida. La fresa: 10 explantos cada 2 semanas. Se obtienen fácilmente 10 millones de plantas en 3-4 meses.
- Multiplicación lenta. El arándano: 3 nuevos explantos cada 6 semanas. Se obtienen unas 20000 plantas al año.
- Multiplicación media. 5 explantos cada 4 semanas. Se obtienen unos 2 millones de plantas en 9 meses.
La forma de controlar al máximo la dominancia apical y promover la ramificación es mediante la aplicación de citoquininas, y si es necesario siempre aportando una determinada concentración menor de auxinas. En la práctica la principal diferencia entre la inducción de yemas adventicias y el incremento de la ramificación exilar es la dosis de citoquininas aportadas, que en este segundo caso es mas baja (por ejemplo una cantidad promedio es de 2 mg/l de Benciladenina)
La mayor limitación intrínseca a este sistema es el exceso de dominancia apical que puedan tener en algunos casos o especies los explantos. Cuando se produce esta dominancia apical cada vez que se produce una nueva rama su meristemo apical inhibe el desarrollo de las yemas inferiores, impidiendo así la ramificación a pesar del aporte de las citoquininas del medio de cultivo. En estos casos debe acudirse a procedimientos que eliminen la nueva dominancia apical cada vez que se produzca, tales como técnicas de “despunte”, “in vitro layering” (peral, Acer, Bétula), o incluso corte individualizado de cada una de las secciones nodales en cada ciclo de multiplicación.
c) Ventajas y desventajas de la multiplicación mediante yemas preexistente
La principal desventaja de este sistema respecto a los anteriores es su menor velocidad de multiplicación, lo que sin embargo no evita poder obtener un número muy elevado de plántulas, y suficientes en la mayoría de las ocasiones, en periodos de entre 6 y 12 meses de cultivo. La gran ventaja es su elevada estabilidad genética, ya que el sistema no altera el tipo de desarrollo del vegetal sino tan sólo promueve la ramificación; no obstante en algunos casos pueden desarrollarse algunos microcallos, que también pueden pasar inadvertidos y causar que al final de la micropropagación haya plantas con mutaciones. Otras limitaciones que se han presentado en algunas ocasiones son la detención del desarrollo de los explantos y la reducción de la longitud internodal de los tallos, lo que requeriría un aporte de giberelinas para facilitar los subcultivos y la obtención final de plántulas; el aporte de giberelinas dificulta el enraizamiento posterior de los explantos, por lo que debe hacerse con sumo cuidado.
Este sistema es el más ampliamente utilizado a nivel comercial, y puede ser aplicado sin problemas a las plantas, sean o no quimeras.
La multiplicación mediante embriogénesis puede ser indirecta o directa en función de que en el explanto se produzca o no un desarrollo de callo previo, si bien en la práctica la embriogénesis directa es mucho mas infrecuente. Por otra parte la multiplicación mediante embriogénesis siempre será mediante estructuras adventicias, ya que no existen embriones preformados más allá de los embriones cigóticos.
Antes de introducirse en la multiplicación mediante embriogénesis conviene recordar algunos conceptos básicos
a) Tipos de explantos
Los embriones no cigóticos se originan a partir de células del esporofito de la planta; la embriogénesis puede producirse a partir de:
Una única célula. Se divide asimétricamente de la misma forma que lo hace un cigoto para dar un embrión, por lo que es una embriogénesis directa
Una agrupación de varias células se divide inicialmente de forma desorganizada, para a continuación organizarse algunas de ellas en una estructura que es el inicio del desarrollo del embrión somático.
b) Fases de la embriogénesis
Cualquiera que sea el inicio de la embriogénesis, se siguen las fases habituales de la misma. Las fases por la que pasa un grupo de células somáticas que sufren un proceso de embriogénesis somática indirecta son:
1. Inducción y proliferación de callo embriogénico. En esta fase el objetivo es la producción de un callo con abundantes masas meristemáticas de las que luego podrán emerger los embriones. Para ello el medio de cultivo debe contener, además de nitratos, amonio como fuente de nitrógeno y también niveles elevados de potasio. En cuanto a reguladores se precisa el aporte de auxinas y citoquininas, preponderando las primeras sobre las segundas.
2. Formación y crecimiento de los embriones. En esta fase es importante reducir hasta casi eliminar el aporte de auxinas, mientras que el de citoquininas se elevará en mucha mayor proporción; también es importante evitar el acumulo de etileno en el interior del recipiente de cultivo, que impide la organización celular necesaria para formar los embriones. Una vez se inicia el desarrollo de los embriones es conveniente aislarlos del callo embriogénico para que puedan formar una estructura independiente. En esta etapa el embrión pasa una serie de fases, típicamente conocidas como fase globular, fase acorazonada y fase torpedo.
3. Maduración de los embriones ya desarrollados. Tras pasar por la fase torpedo el embrión empieza a madurar, es decir, pierde agua para dar lugar a una estructura vegetal con una proporción de agua mucho menor que la de una planta en desarrollo, que es el embrión maduro, y que se puede conservar con relativa facilidad en ese estado durante un tiempo prolongado. Para favorecer la maduración de los embriones se ha propuesto la aplicación de ácido abscísico en el medio de cultivo, que se basa en el aumento natural en los niveles de dicho regulador al final del proceso de maduración de las semillas en la planta madre y en su relación con la resistencia de la semilla a la desecación. Alternativamente, los embriones ya desarrollados se pueden someter a un lecho de aire estéril seco que los vaya desecando poco a poco; esto puede hacerse dejándolos durante 12-24h sobre placas Petri abiertas en el interior de la cabina de flujo laminar en funcionamiento.
4. Germinación. Los embriones desecados pueden volver a hidratarse y germinar si se colocan en las condiciones adecuadas de humedad y temperatura, siempre que no se encuentren en dormición. Los embriones también se pueden encapsular de forma que constituyan una semilla artificial que pueda conservarse o sembrarse de forma equivalente a como se hace con las semillas típicas de la especie. En esta encapsulación se pueden añadir compuestos que faciliten la germinación, incluyendo aspectos como la utilización de fungicidas en su pildoración.
Los reguladores que se utilizan para producir una embriogénesis indirecta coinciden en general con los utilizados para producir una organogénesis indirecta; las diferencias principales se basan en las cantidades y proporciones relativas de auxinas y citoquininas utilizadas. Además, estos requerimientos difieren según la especie a cultivar, si bien existen unas tendencias comunes según la planta sea monocotiledónea o dicotiledónea. Las pautas generales para organogénesis o embriogénesis indirecta de dicotiledóneas son las comparadas en la tabla.
c) Patrón de desarrollo del embrión somático, eficiencia del sistema y subcultivos
Es importante reconocer al embrión al inicio de su desarrollo sobre las masas meristemáticas del callo embrionario, de forma que podamos aislarlo, ya que en caso contrario su desarrollo podría quedar comprometido por la alta tasa de división celular del resto de las células meristemáticas de este callo. Para ello es importante su reconocimiento mediante:
- Caracteres morfológicos.
- Cutinización de la superficie de las células que inician el desarrollo embrionario.
- Incremento de nutrientes que se acumulan como sustancias de reserva o estructuras celulares, dando a las células un aspecto denso y opaco.
Posteriormente se podrá realizar un seguimiento visual del desarrollo embrionario, según las tres fases mencionadas anteriormente y características de las especies dicotiledóneas; en las monocotiledóneas el proceso es paralelo pero algo diferente.
Otro aspecto a considerar es que si bien el embrión somático pasa por los mismos estados de desarrollo que el embrión cigótico, su diferente origen permite algunas variaciones. Entre ellas, e incluso si el embrión se inicia a partir de una única célula, a lo largo de su desarrollo las células que se producen pueden dividirse según diferentes opciones y dar lugar finalmente a un embrión originado de diferentes formas.
En cuanto al número de subcultivos que pueden realizarse durante la fase de multiplicación por embriogénesis indirecta, éstos corresponden exclusivamente a la primera fase, es decir a la proliferación de callo embriogénico. Este callo, a semejanza con el callo organogénico, puede ser fragmentado y subcultivado de forma que se incremente el número de callos embriogénicos, con lo que al final de la fase de multiplicación se puede tener un número indefinido de éstos. Posteriormente en cada callo embriogénico se induciría la formación de varios embriones al cambiar de medio de cultivo. Sin embargo el número de subcultivos posibles en la práctica no es ilimitado, ya que por encima de un determinado número se ha observado que los callos van perdiendo capacidad embriogénica. Este número depende de la concentración de reguladores de crecimiento en el medio de cultivo y de la especie cultivada, y se han formulado tres hipótesis explicativas, la hipótesis fisiológica, la hipótesis genética y la hipótesis competitiva que combina las dos anteriores.
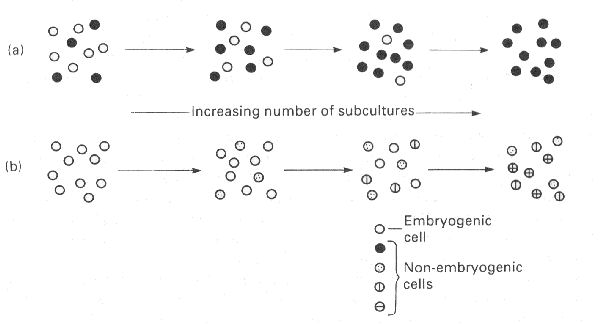
Esquematización de la hipótesis competitiva.
d) Anomalías del desarrollo embrionario
El desarrollo embrionario no está exento de sufrir anomalías que impidan o dificulten su germinación o la obtención de nuevas plántulas completas. Estas anomalías también se producen de hecho en los embriones cigóticos formados mediante reproducción sexual convencional, y serían responsables de diversos problemas bien identificados que limitan la germinación de las semillas y su posterior viabilidad. Un proceso de embriogénesis somática es por ejemplo el detallado para zanahoria; las principales anomalías identificadas son la formación de embriones con más de un ápice o sin el ápice bien definido, embriones con más de un extremo radicular, embriones con más de dos cotiledones, embriones fusionados que comparten algún grupo de células, o embriones que inician su formación sobre otro embrión previamente formado y en estado de crecimiento. Esta última anomalía de la embriogénesis somática es relativamente frecuente y difícil de detener, y se conoce como embriogénesis recurrente; al producirse de forma continuada una y otra vez es un problema grave para la obtención final de embriones maduros.
e) Ventajas y desventajas de la embriogénesis somática.
Aplicaciones. El sistema se puede utilizar para todo tipo de plantas excepto las que sean quimeras o presenten un patrón de desarrollo que simule una quimera. Es un procedimiento muy valorado en los programas de mejora, ya que los callos son un tipo de tejido muy favorable para inducir mutaciones o seleccionar plantas con caracteres concretos, y a partir de ellos la producción de numerosos individuos mediante embriogénesis es muy rápida y bastante estable.
Al implicar la obtención inicial de callo en la gran mayoría de las ocasiones existe una cierta prevención a utilizar este método, sin embargo la aparición de plantas con mutaciones es mucho menos frecuente utilizando embriogénesis somática indirecta que mediante organogénesis indirecta. La razón se atribuye a los mayores requerimientos de integridad y de organización celular que deben mantener las células para dar lugar a un embrión, frente a los requeridos para iniciar una yema adventicia; gracias a ello es posible en muchas ocasiones micropropagar una planta mediante embriogénesis somática aunque sea una vía indirecta. Los embriones producidos serán clónicos.
Este procedimiento, además de cómo una de las opciones de micropropagación, se ha utilizado como un método de saneamiento de plantas portadoras de virus, como es el caso típico de los cítricos. Mediante la formación de embriones somáticos en explantos formados por la nucela de dichos cítricos pueden regenerarse plantas sanas.
Finalmente, podemos observar algunas de las fases de la producción de embriones somáticos y su posterior germinación, así como un esquema con un caso práctico de embriogénesis somática a partir de un cultivo de callo embriogénico y, alternativamente, a partir de un cultivo de células en suspensión.